"Because the world is round it turns me on.
Because the wind is high it blows my mind.
Because the sky is blue it makes me cry."
Because by John Lennon and Paul McCartney, 1969
Intrepid explorer and self-taught geologist John Wesley Powell, during his historic expedition down the Green and Colorado Rivers in 1869, astutely recognized the Great Unconformity but at the time was incapable of appreciating its enormous temporal magnitude and scientific significance. Located globally, it's a contact between a Precambrian basement of igneous and metamorphic rock and a cover of Phanerozoic sedimentary strata - both of which can vary in age.
In side-canyon Blacktail at the bottom of the Grand Canyon, it's ~1.2 billion years in duration - almost one-fourth the age of the Earth. You can drag your fingers across the gap of missing strata and time, as Powell assuredly did and every geologist and river runner that followed. At Rattlesnake Mountain anticline in northwestern Wyoming, it's over 2 billion years - nearly twice as long and almost half the age of Earth - and you can drive right up to it!
 |
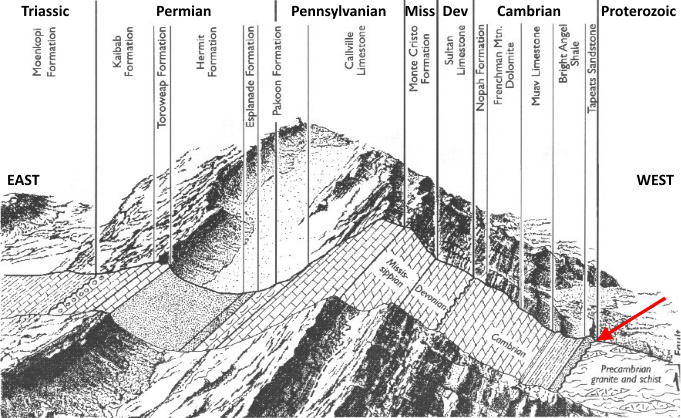
The Great Unconformity of Shoshone Canyon
Flat-lying, brown, maroon and white Cambrian-age Flathead Sandstone overlies tilted, pink and black crystalline rocks of the Late Archean Wyoming Craton. An early stream channel on land delivers arkosic sands to the Great Unconformity about to form.The vertical striations are from construction of the roadcut. |
How did this deeply buried geologic feature form? Why is it found worldwide in so many disparate locations and at so many inexplicable elevations? Why is it so much longer in duration at Rattlesnake anticline than in Arizona's Grand Canyon and inclined on the landscape? Let's investigate.
ABOUT THIS POST
This is my fourth post on the Great Unconformity. In Part I (here), I discussed its genesis in Grand Canyon. In Part II (here), I offered a more global interpretation of its presence. In Part III (here), I discussed the time-gap at Baker's Bridge in southwest Colorado and its theorized significance to biological evolution.
In this Part IV, my daughter and science teacher Julia Share, geologist Wayne Ranney (here) and I, while geo-journeying through western Wyoming, visited the celebrated contact. Herein, I discuss the Great Unconformity's relationship to the Archean cratonic evolution of the Wyoming Province and its curious Laramide disposition on the landscape.
Italicized terms are defined, important names and locations are highlighted in boldface where first mentioned, and global coordinates of various sites and locales are provided that can be pasted into an on-line mapping program such as Google Earth, so you can "Go there."
 |
Promontory Point, Utah
After converging in Salt Lake City and heading into Wyoming, we stopped some 60 km northwest of of the city where the Union and Central Pacific Railroads met and celebrated the event with the Golden Spike Ceremony in May, 1869. We raced to visit the National Historic Park before closing, but were too late. Instead, we got a wonderful private tour of the site by the park ranger. Go there: 41°37'4.78"N, 112°33'3.44"W |
WHERE ARE WE?
It's safe to say that most passersby aren't aware of its presence, let alone know of its existence or comprehend its scientific significance. But for geologists, who cherish and revere the iconic geologic feature, it's difficult to miss. For them, with cameras ready, seeing it is a source of anticipation and excitement and requires paying homage, which we assuredly did.
 |
Missing Time in the Excavated South Face of Rattlesnake Mountain
The Great Unconformity is exposed in the roadcut on both sides of the highway before the first tunnel west of Cody. Buffalo Bill Dam and Reservoir appear ~1.3 km after the third tunnel. Below the tilted contact spans rocks of Wyoming's Archean basement through which the tunnel has been cut. Above the contact lies the dipping base of the Paleozoic sedimentary column that was deposited on Laurentia's passive margin. Go there: 44°30'33.32"N, 109°10'22.11"W |
WHAT'S AN UNCONFORMITY?
The norm rather than the exception is that successive strata do not always form in an uninterrupted manner. Interruptions or cessations in deposition result in a geologic contact or inter-stratal boundary between rock layers. It's a "temporal gap" between an older surface that has been subjected to erosion and an overlying, younger one that experienced a hiatus in deposition. Thus, an unconformity is a buried surface of missing strata, unseen until exposed.
They vary in type depending on composition of the juxtaposed layers and occur globally on our ever-evolving planet. They can be thousands or even millions of years in duration, if not more. In Grand Canyon, there are so many "time-gaps" (at least 14 disconformities) in the mile-high Paleozoic record that the sum of missing time is greater than that preserved, although there is one 'great' unconformity that exceeds all others.
 |
Three Flavors of Unconformities
A disconformity of irregular topography forms between sedimentary rocks. An angular conformity forms when the underlying rock layer is tilted or contorted. The Tapeats overlying the Supergroup is an example. Nonconformities such as the Great Unconformity occur when the underlying layer is either metamorphic or igneous rock that has been eroded before being covered by sediments. |
WHAT'S THE GREAT UNCONFORMITY?
It's a profound break in the Earth's rock record. Typically, it resides above a denuded (eroded and exposed) Precambrian basement of igneous and metamorphic rocks and below flat-lying sedimentary rocks deposited at sea level during the Paleozoic (often at the base of the Cambrian system) or Mesozoic. The temporal magnitude of the "buried erosion surface" is governed by the age of the juxtaposed strata, which can vary greatly.
Highly recognizable and a composite of many events that occurred, it's found on daughter continents that fragmented from the Late Proterozoic supercontinent of Rodinia and, "rifting and drifting", were tectonically carried afar across the globe. Yet in spite of its ubiquity, it's one of the Earth's most enigmatic features and great challenge for geoscientists to decipher what happened geologically, climatologically and biologically during the immense interval of its formation.
 |
The Great Unconformity of the Grand Canyon
Easily seen from the canyon's South Rim, the high-points of Late Proterozoic-age Vishnu Schist were rocky islands poking out of a Cambrian sea before being blanketed with the Middle Cambrian Tapeats Sandstone that formed the Great Unconformity. Go there: 36° 6'7.93"N, 112° 7'58.48"W |
MISSING TIME WITHIN THE GRANITE GORGE Where is it found? One of the Great Unconformity's most celebrated and visited localities is at the bottom of northern Arizona's Grand Canyon within the oldest section of Granite Gorge. In an unsuspecting slot canyon called Blacktail that enters the Colorado River at grade, the contact comes down from its lofty perch along the walls of the gorge and lies only a few feet above Blacktail's ever-changing, flash flood-delivered, cobble-strewn floor.
The contact lies between ~1.75 Ga Vishnu Schist - the collective name for the canyon's Early Proterozoic-age igneous and metamorphosed basement rocks - and the overlying ~500 Ma Cambrian-age Tapeats Sandstone. The missing ~1.25 billion years is almost 25% of the age of our ~4.5 billion year old planet!
 |
Blacktail Canyon
With walls comprised of slabs of Tapeats Sandstone, the Great Unconformity resides just above the side canyon's river cobbled floor. Go there: 36°14'24.43"N, 112°28'21.10"W |
You can touch the contact as John Wesley Powell must have and stand in contemplative awe of its inconceivably immense magnitude and ponder the global events that may have happened during its formation. Like missing pages of a history book, those secrets are slowing being revealed. That's been the challenge of the Great Unconformity since its discovery!
 |
Geologist Wayne Ranney Touches Blacktail's Great Unconformity
A heat and pressure-gnarled Vishnu basement lies beneath earliest transgressing beach sands of the Tapeats. Notice the chunks of metamorphosed granite and schist that were kicked up by turbulent waves of a rising sea a half-billion years ago and incorporated into the sandy matrix. Wayne has explained the Great Unconformity's mystery and geology to countless throngs of geologists and river runners. |
Blacktail is flocked to as if a hallowed shrine. Standing in its quiet shadows and soft echoes over a decade ago, geologist Wayne Ranney read a moving poem entitled "The Great Unconformity", which he does on every river trip in Grand Canyon. This is the poem's elegant introduction. You can listen to Wayne read it here.
"It is time
Touch it
Trail your fingers over a billion years
Etched in the red rock
There is no monument, no temple more sacred
Than this monumental temple to the whole wide world
Seared this great chasm of whispering, eroded eons
With majesty beyond comprehension"
by Charlotte Graham-Clark
IN EASTERN GRAND CANYON
Another spectacular place to view the Great Unconformity is from Lipan Point and nearby Desert View on the South Rim of eastern Grand Canyon. It's also where the spectacular Grand Canyon Supergroup is on display. The massive, gently tilted, two-suite complex includes the Mesoproterozoic Unkar and Neoproterozoic Chuar Groups of largely sedimentary and lesser igneous rocks.
They were deposited within intracratonic basins (versus deep oceanic basins) during the final completion and fragmentation, respectively, of supercontinent Rodinia - the latter thought to have occurred just prior to the onset of "Snowball Earth" global glaciation. The geologic history-revealing Supergroup formed during the interval of the Great Unconformity below the Tapeats and above the Vishnu basement.
 |
The Great Unconformity of the Grand Canyon
The Unkar Group records the arrival of island arcs and continent-continent collisions that completed the formation of Rodinia during the Grenville orogeny. The Chuar Group was deposited in an intracratonic marine basin(s), when extension rifted apart the supercontinent prior to global glaciations of the Cryogenian period at the end of the Proterozoic. The group contains molecular and micro-fossiliferous clues to early eurkaryotic life and biological diversification before the advent of complex multi-cellular organisms. From Karlstrom and Timmons, 2012
|
FORTUITOUS EXPOSURE
Had it not been for extension in the Proterozoic, faulted crustal blocks of the Supergroup in the Grand Canyon would never have downdropped and tilted that protected them from erosion. Without its fortuitous tectonic preservation and subsequent exposure by action of the Colorado River, we would know far less about global events that happened at the time.
Viewed from atop Desert View Tower, tilted and faulted Unkar strata contact the flat-lying Tapeats at an angular unconformity (arrows). The size of the 'lesser' Great Unconformity that formed varies in duration contingent on the age of the particular stratum that lies below the contact and grows progressively longer down-sequence beginning with dark, interbedded Cardenas Basalt followed by the purplish, sedimentary Nankoweap Formation and so on.
 |
The Great Unconformity in Eastern Grand Canyon
Go there: 36° 6'20.34"N, 111°52'4.61"W |
OFF THE COLORADO PLATEAU IN SOUTHERN NEVADA
West of Grand Canyon and Lake Mead and immediately east of Las Vegas, the Great Unconformity is exposed on the west-facing slope of Frenchman Mountain. A nondescript pullout on Lake Mead Boulevard (SR147) and a short hike will bring you to the contact where interpretative signage marks the spot.
Tilted about 50° synonymous with the strata of the entire edifice, the temporal gap (red arrow) of the Great Unconformity lies between Cambrian Tapeats Sandstone and underlying Proterozoic-age Vishnu crystalline basement rocks. The entire assemblage mimics that of the Grand Canyon and angled in its disposition on the landscape! How did this happen?
 |
West-East Cross-Sectional View of B&R Tilted Frenchman Mountain
Modified from geoscience/unlv.edu/pub/rowland/virtual/geology.html
|
In every locale of the Great Unconformity, its geologic story involves a discussion of its duration, how its juxtaposed layers formed, how it was tectonically delivered to the locale, how it achieved its elevation and orientation on the landscape, and how the deeply buried contact became exposed.
The hint to the provenance of the tilted, NS-striking, small range of Frenchman and Sunrise Mountains is in its position between Lake Mead Basin on the east and Las Vegas Valley on the west. Collectively, they are a small part of the distinctive landscape of the Basin and Range Province that spans much of inland Western U.S. and northwestern Mexico. Its signature landform are alternating, flat sediment-filled valleys and NS elongate, fault-bound mountain ranges, one after the other, all essentially on strike.
 |
Frenchman and Sunrise Mountains from the East
Traveling west on Lake Mead Boulevard (SR147) across Lake Mead basin, the dark dacitic knob of Lava Butte is in the foreground (and others unseen) form a lava field that emplaced some 13 myr in the mid-Miocene, far younger than the Paleozoic rocks of Frenchman and Sunrise to the north (right) in the distance. Extension transported the Frenchman-Sunrise range and produced associated magmatism across the landscape. Las Vegas Valley lies beyond the range. Go there: 36° 9'13.21"N, 114°53'21.00"W
|
The demise of the Farallon plate and arrival of the spreading center between the Pacific and North American plates in association with collapse of the Sevier Highlands is responsible for pervasive extension of the crust in the province. In this manner, Frenchman and Sunrise originated some 50 to 65 km from the east, possibly associated with the Gold Butte block located between Lake Mead and the Colorado Plateau.
Following Sevier phase uplift related to descent of the Farallon (explained below), the Frenchman-Sunrise allochthonous block (transported crust) of Paleozoic and Proterozoic strata (same as the Grand Canyon) with the Great Unconformity sandwiched in between, was transported in the Miocene along detachment (low-angle extensional) and strike-slip (side-to-side) faults to its present location.
 |
Wayne Ranney Identifies the Great Unconformity on Frenchman
Above the extensionally tilted contact lies the basalmost bed of Cambrian-age Tapeats Sandstone and below, pinkish metamorphosed rocks of Proterozoic-age gneiss and schist. Go there: 36°11'57.31"N, 114°59'48.36"W
|
MISSING TIME IN COLORADO
Here's another example. East of the Colorado Plateau and at an elevation of ~7,500 feet, it's exposed within the towering Painted Wall of Southwest Colorado's Black Canyon of the Gunnison. Only there, without any interposed Paleozoic strata, ~1.5 billion years of time are missing between Paleo- to Mesoproterozoic rocks of Black Canyon gneiss and overlying Entrada Sandstone. What's the genetic explanation there?
The Pennsylvanian-Permian Ancestral Rocky Mountain Range - long-gone predecessor of the modern Rockies - was eroded and buried by the Triassic. At Gunnison, on a planed-off remnant of the highlands, the Great Unconformity formed when a 'sand sea' blanketed it in the Jurassic. The gneiss-blanketing erg formed on the southern terrestrial margin of the Sundance Sea, which was a short-lived, epeiric (shallow continental versus deep marine) embayment of the early Pacific Ocean in western North America.
 |
Facing North Toward the Painted Wall in Colorado's Black Canyon of the Gunnison
Go there: 38°34'32.71"N, 107°45'6.94"W |
AROUND THE ADIRONDACKS OF NEW YORK STATE
It's located across North America on the periphery of the Adirondack Mountains in northern New York State's Saint Lawrence and Lake Champlain Lowlands and in the northern Mohawk River Valley. At each location, the contact lurks almost at sea level between the Middle Cambrian Potsdam Sandstone and underlying Mesoproterozoic Grenvillian gneiss that emplaced and metamorphosed as Pangaea completed unification during the protracted Grenville orogeny.
 |
Ausable Chasm in Northeastern New York State
The 1.5 mile-long sandstone gorge of Potsdam Sandstone is a Tapeats-equivalent. The Ausable River runs through it and empties into Lake Champlain in northeastern New York State near the small town of Keeseville. Unseen at this locale, the Great Unconformity lies buried a few hundred feet beneath the basal Sauk sequence on the northeast side of the Adirondack dome. From Wikimedia Commons and Alan Prusila |
The Potsdam Sandstone was deposited during the global Salk transgression on proto-North America's eastern margin, as was beach sand of its Tapeats geo-equivalent on the western margin. Only partially understood, ongoing Cretaceous uplift of the Adirondack dome - creating "new mountains from old" - unroofed the Potsdam, while uplifting the Great Unconformity and underlying gneiss.
In addition, inliers (isolated, older blocks of rock) containing the Great Unconformity lie buried - in the collision between Laurentia and Gondwana that completed the formation of Pangaea - beneath the Appalachian Mountain Range along the continental margin of eastern North America. Following fragmentation of the late Paleozoic supercontinent in the Mesozoic that severed the Central Pangaean Range into the Appalachians and Scandinavian Caledonides, the Great Unconformity was tectonically delivered to Norway, Sweden, Estonia and Finland via microcontinent Baltica across the widening Atlantic Ocean.
 |
Great Unconformity of the Saint Lawrence Lowlands
Here it is again northwest of the Adirondack dome. Go there: 44°28'2.29"N, 75°45'47.64"W. It's also in the Lake Champlain Lowlands here: 3°25'44.05"N, 73°28'10.22"W |
IN THE TETONS
Back in Wyoming, the geologic contact (red arrow) is at nearly 12,610 feet on the summit of Mt. Moran in the Tetons. Vertical en bloc uplift of the Teton range along 40 mile-long Teton fault, elevated the GU beneath a tiny, remnant cap of Cambrian Flathead Sandstone overlying the range's ~2.7 billion year old gneiss of Archean-age - a time-gap of over 2 billion years!
That places it 6,000 feet above Jackson Hole's valley floor and ~20,000 feet above the sediment-buried unconformity beneath the crustal block's downdropped basin. The magnitude of the time-gap in Wyoming, in contrast to that of the Grand Canyon, is related to the extreme age of its Archean-age basement.
 |
Mt. Moran and Jackson Lake in Grand Teton National Park
Go there: 43°50'1.08"N, 110°46'36.77"W |
ON DISPERSED RODINIAN DAUGHTER CONTINENTS
Tectonically severed remnants of the Great Unconformity were transported across the globe following the diachronous, protracted fragmentation of supercontinent Rodinia beginning in the latest Proterozoic. As a result, exposed by erosion and uplift, it's found on "rifted and drifted" continents in places across the globe.
Examples are South Africa's Table Mountain (on the Kaapvaal craton), at Huaibei (on the North China craton) and Wadi Rum near Jordan's Petra (on the Arabian-Nubian Shield). At each location, the Great Unconformity has a similar and highly recognizable appearance.
 |
The Great Unconformity at Petra
Courtesy of Geologist Wayne Ranney
|
AT SHOSHONE CANYON IN WYOMING
At Grand Canyon's Blacktail, it saw the light of day facilitated by en masse uplift of the Colorado Plateau and exposure by the Colorado River and its system of tributaries. In contrast at Wyoming's Rattlesnake Mountain where we're pulled over, it too was uplifted and then subsequently wrinkled on the landscape during subduction of the Farallon oceanic plate. It took construction of a new highway facilitated by canyon-forming fluvial incision to expose the buried contact.
 |
The Great Unconformity of Shoshone Canyon
Our view is to the north across the highway in a roadcut excavated into the south face of Rattlesnake Mountain. Cody lies a few minutes to the east and the tunnel entrance is just to the left. The vertical drill lines were created during construction of the highway. |
ARCHEAN NORTH AMERICA IN THE BEGINNING
In order to more fully appreciate the evolution and duration of the GU at the Rattlesnake Mountain anticline, let's briefly summarize the evolution of North America. To the best of our understanding (or what we think we understand), no large continents existed in the early Archean Eon (4,000 to 2,500 Ma), almost 4 billion years ago.
But, by mid-eon around 3 Ga (although some say much later), the Earth's crust had cooled enough to initiate surface plate rigidity and mobility, mantle recycling and subduction-related processes. As a result, a global mosaic of cratons (stable crustal blocks) and terranes (independent blocks with distinctive histories and composition) began to tectonically collide and accrete (weld together), building a continent of increasingly larger coalesced landmasses.
 |
Archean and Proterozoic Foundation of Laurentia
The core of the North American continent (Canadian shield) formed in the Proterozoic by plate collisions of Archean continents and smaller Archean continental fragments such as the Wyoming Province. The ultimate orogenetic collision provided Laurentia with the Grenville Province that brought together the known land masses of the world to assemble the supercontinent of Rodinia. Whitmeyer and Karlstrom, 2007. |
In Canada, Archean age blocks constitute geologic provinces (sizable areas with common structure, composition and genetic origin) with names like Slave, Superior, Trans-Hudson, Rae, Hearne and Wyoming in the US. The cratonic jigsaw puzzle - referred to as the Laurentian craton or just Laurentia - has remained unified within a succession of supercontinents and continents that followed that have cyclically assembled, dissociated and reassembled.
THE ARCHEAN WYOMING PROVINCE
Long considered one of the oldest terranes in North America, the Wyoming Province coalesced from even older amalgamated Archean crustal blocks that accreted to Laurentia's southwestern margin and parts of adjacent states around 1.9 billion years ago. They're distinguished geochemically, petrologically and structurally from older Archean provinces.
Collectively, they form the Canadian Shield, the Laurentian core of North America, which is a large exposed area of stable Precambrian crystalline basement (foundation of igneous and high-grade metamorphic rocks). Wyoming's Late Archean rocks also form a portion of the complex basement of the Cordilleran system of mountain ranges in western North America. It's dominated by 2.8-3.0 Ga gneissic rocks intruded by 2.5-2.7 Ga granitic plutons and overlain by supracrustal rocks of similar age.
THREE WYOMING SUB-PROVINCES
The Wyoming Province can be subdivided into three major subprovinces of Archean crust. From oldest to youngest and basically NW to SE, they are: the Montana Metasedimentary Province (MMT), the Beartooth-Bighorn Magmatic Zone (BBMZ) and a collage of Southern Accreted Terranes (SAT). South of the Wyoming Province (below the map 'teethed' tectonic collision zone below) are accreted Paleoproterozoic orogenic belts of the Cheyenne Belt (CB), Great Falls Tectonic Zone (GFTZ), Farmington Zone (FZ) and the Trans-Hudson Orogen (THO) of reworked Archean crust.
The BBMZ in Northwest Wyoming and southern Montana contains largely metamorphosed plutonic (igneous solidified at depth) rocks that cratonized (formed) ~2.8 Ga and welded to Laurentia ~2.7 to 2.5 (1.8 and 1.7) Ga. It forms the deeply buried crystalline basement of Bighorn Basin, the exposed walls of Shoshone Canyon and constitutes the lower component of the Great Unconformity in Northwest Wyoming.
 |
Ancient Crystalline Rocks of the Beartooth–Bighorn Magmatic Zone
Much is unknown and conjectured about Wyoming's Archean rocks. In the walls of Shoshone Canyon are ~2.8-3 Ga metamorphosed plutonic rocks that are largely granitoids such as tonalites and granodiorites that are closely related to 'true' granites. Even older Archean gneisses (>3 GA) are present that are interpreted as inherited cores. |
TECTONIC GROWTH OF OF THE SOUTHWEST
Things didn't end in the Archean. In the early and middle Proterozoic (2,500 to 541 Ma), convergent orogenic tectonism continued with tectonic acquisition of the Mojave, Yavapai and Mazatzal provinces. Building mountains and adding crust, they constitute a succession volcanic and plutonic rocks that formed when intra-oceanic volcanic arcs and adjacent ocean basins collided and sutured to Wyoming's southwest and southern margins.
The southwest of proto-North America was under progressive Proterozoic construction that ended in the Middle Proterozoic with the protracted, multi-stage Grenville orogeny. The Grenville province likely formed when the Amazon craton of South America collided with the southern margin of Laurentia. Characterized by large-scale magmatism and extensive deformation, it represented the final assembly of all known global landmasses into supercontinent Rodinia.
 |
Map of Precambrian Provinces in Western North America
The Late Archean cratonic Wyoming Province (encircled) lies juxtaposed to Proterozoic rocks of the Mojave and Yavapai rocks on the south with Mazatzal and Grenville completely the Southwest's assemblage to the southeast. Purple areas represent exposures of Precambrian rocks among a thick cover of Phanerozoic sedimentary rocks. Note the location of Cody just east of the Great Unconformity exposure. Modified from Foster, 20015.
|
THE BIG PICTURE
The Laurentian core now possessed a basement crust with both Archean (in the north) and Proterozoic (in the south) affinities. It's the foundation of the Great Unconformity, yet to form. All that was needed - an incredibly long temporal requirement - was uplift and erosive denudation of the basement followed by the deposition of sedimentary overburden in the Phanerozoic, typically Cambrian.
The 'waiting period' between the two components represents the Great Unconformity's profoundly long temporal gap! Thus, in Arizona's Grand Canyon it resides on Yavapai-derived, Late Proterozoic Vishnu crust, while in Wyoming, the contact lies on Archean crust of the Beartooth-Bighorn Magmatic Zone.
 |
| Wayne Photographs the Great Unconformity on the South Side of the Highway |
TRANSGRESSING SEAS AND OCEAN-DERIVED SANDS
The Great Unconformity formed when rapid rising seas in early to mid-Cambrian time gradually flooded the western margin and accessible interiors of Laurentia, the term for the ancient and stable, continental core of North America. The transgressive process was facilitated by post-rift, thermal subsidence of nascent continental margins and climate change that brought about near-global eustasy (rising levels of the sea).
In the Cambrian, the Sauk global marine transgression delivered sedimentary sequences over the Laurentian crystalline basement of foundation rocks that are of Archean age to the north and Proterozoic age on the south. It blanketed the coastal margins of long-gone supercontinent Rodinia's rifted daughter continents with a layered mix of sandstone, shale and limestone.
 |
Transgressing Early Cambrian Sea along Western Laurentia
Beginning in the Cambrian and continuing through most of the Paleozoic, rising seas flooded newly rifted margins of Laurentia and blanketed denuded crystalline basement rocks. Red dots indicate locations of the Great Unconformity discussed in this post. Furthest west, the Frenchman GU formed before the others as the sea flooded the continent. The idealized river at the Grand Canyon GU is not the modern Colorado. Farallon contractional subduction uplifted every GU, whereas Basin and Range extension at Frenchman lowered it somewhat. Modified from Colorado Geosystems of Ron Blakey. |
First to form was the Tapeats (and its equivalents such as the Flathead). It's the basalmost sequence of Grand Canyon's mile-high column of flat-lying, Paleozoic rocks that culminated in the formation of the Great Unconformity.
At that time, western Wyoming and the region of the future Rattlesnake anticline were along the Cambrian coastline. It remained as such though the majority of the Paleozoic, until a number of incoming exotic terranes, island arcs and oceanic plates added to the crustal mass of western North America and 'drove' Wyoming further inland beginning in the Mesozoic.
SOME IMPORTANT FARALLON STUFF
The GU varies in elevation and topographic setting as a result of tectonic changes western North America experienced in the Mesozoic and Cenozoic. It's largely attributable to SW-NE oblique subduction of the oceanic Farallon plate beneath the western edge of the west-migrating continental North American plate.
It's also related to the 'interaction' of the Farallon with Laurentia's Precambrian basement, prone-to-fail "zones of weakness" inherited from past tectonic events AND overlying Phanerozoic sedimentary cover.
Farallon descent began in the latest Jurassic and terminated in the Oligocene. It was the longest-lived plate (over 150 myr) and the largest (most of the proto-Pacific basin). It was also the most important for the landscape changes produced AND the daughter plates it spawned. Examples are the subducting Juan de Fuca in the Pacific Northwest, Cocos west of Mexico and Nazca along western South America.
 |
Early Fragmentation of Pangaea in the Late Jurassic
The nascent continents of the Atlantic realm have formed across a newly formed, widening ocean. Driven by its expansion, the Farallon plate has begun to subduct beneath the western margin of Laurussia, the late Paleozoic core of North America. Its descent will progress in two phases that reflect a progressively shallower angle of descent during the Sevier and Laramide orogenies. Modified from Ron Blakey's Colorado Geosystems Inc. |
TWO OVERLAPPING PHASES OF THE TWO-PLATE COLLISION
Collision of the Farallon plate and a number of 'suspected' (not fully resolved), 'exotic' terranes (landmasses made-elsewhere) with the North American plate produced a long line of continental volcanic arcs and crustal (lithospheric) compression in western North America. Its descent initiated at a normally high-angle that shallowed over time and produced a change in the "style of deformation" during a bi-phasic orogeny (mountain-building event) that overlapped in time and space.
The Sevier ororgenic phase...
From latest Jurassic to Middle Eocene, Sevier orogenics did not involve the continent's Precambrian basement. It's referred to as "thin-skinned" deformation of the sedimentary cover that accumulated along western proto-North America's passive margin during the Sauk marine transgression (one of six during the Phanerozoic) that followed in many areas of North America. Part of the cover has decoupled from the basement. The Rocky Mountains provide the best example of this type of tectonics.
Compressive deformation resulted in progressive west-to-east stacking cratonward (continent-ward) in the foreland region (in front of the mountain-belt) of the Farallon subduction zone. Called the Sevier fold-and-thrust belt, the topographically rugged region is largely in Utah and Nevada along the majority of the (then) continental margin from Canada into Mexico. The belt has been largely fragmented by crustal extension in the Cenozoic following and due to demise of the Farallon slab.
 |
Idealized Schematic of Two-Phase Farallon Plate Kinematics
a) Normal, high-angle, thin-skinned Farallon descent during the Sevier phase of orogenics
b) Low-angle, thick-skinned descent of the overlapping Laramide phase with involvement of both Precambrian basement and overlying Phanerozoic sedimentary rocks |
The Laramide orogenic phase...
Often confused with Sevier deformation (I know I was), it occurred from Late Cretaceous through mid-Eocene time (~80 to 45 myr) and continued crustal-shortening but even further east of the plate margin. Driven west at a higher rate of convergence during Pangaea fragmentation, the Farallon assumed a shallower "flat-slab" angle of descent with greater compressive stress.
The "thick-skinned" event includes deformation of the basement AND overlying marine Phanerozoic sequences further into the interior. The cover has remained welded to the basement and includes a compressional reactivation of inherited basement faults.
A source of some confusion, the two styles of deformation - thin and thick-skinned - are not mutually exclusive, that is, they may occur coevally. Laramide tectonics uplifted and overprinted portions of the Sevier belt and foreland in central and eastern Utah, western Colorado and most of Wyoming. It produced a pattern of discontinuous, NW to SE "basement-cored" arching uplifts and intermontane basins across the landscape.
 |
Generalized Tectonic Map of western US in the Paleocene (~60 Ma)
The Laramide belt developed far into the continental interior during the second phase of flat-slab subduction in the Late Cretaceous to Paleogene flat-slab subduction that was oriented SW-NE, oblique to the plate margin. The belt is characterized by an anastomosing network of overall NW-SE to N-S, to locally W-E oriented, fault-bounded, Precambrian basement-cored arches, separated by broad basins Modified from Weil, 2016 |
A few observations regarding the map...
• Basement-cored arches (map above) share a similar orientation that is consistent with oblique Farallon compression delivered from the southwest.
• Laramide kinematics did not appreciably deform the Colorado Plateau but contributed to ~2 km of en masse uplift of the intact crustal block's Precambrian basement and overlying strata.
• It elevated the Grand Canyon's Great Unconformity, while promoting an unroofing (removal of successive layers of strata) across the Colorado Plateau while enabling carving of the canyon that exposed the temporal gap.
BASEMENT-CORED UPLIFTS AND ADJACENT INTERMONTANE BASINS
Bighorn Basin and surrounding uplifts in Northwest Wyoming are archetype examples of Laramide landforms and thick-skinned foreland deformation that developed far inland from the plate margin in the Laramide belt that spans the region from southern Montana to New Mexico.
They are characterized by fault-bound, basement-cored arches with steep marginal thrust faults that are bordered by flanking fold systems and separated by broad basins with an overall NW-SE structural grain. Laramide deformation produced most of the across the reactivated craton of western North America and corresponds roughly with the eastern front of Sevier deformation.
 |
Simplified Cross-Sectional Tectonic Map of the Cordilleran System in the Paleogene (~60-50 Ma)
The Laramide foreland belt developed flat-slab subduction and partly overlapped later phases of Sevier thin-skin thrusting of the fold-thrust belt. Modified from Yonkee, 2017.
|
A ROAD TRIP THROUGH "CODY COUNTRY"
Let's take a short drive west from the town of Cody to Buffalo Bill Reservoir and Dam in order to experience the tectonic history and unique locale of the Great Unconformity. To get there, we'll pass through a massive, V-shaped gap between Rattlesnake Mountain on the north and Cedar Mountain on the south. Along the way, we'll visit the exposure of the unconformity.
At 1,500 m above sea level, Cody (pop. ~10,000) lies along the Shoshone River on the western edge of Bighorn Basin. A little larger than Vermont at ~10,400 sq mi in northwestern Wyoming and some of south-central Montana, it's an asymmetric oval in shape that feels like a broad, oval-shaped valley. It's one of many large synclinal downwarped depressions formed by Laramide compressional deformation across Wyoming and neighboring states. As such, it's sediment-filled with a Precambrian basement and structurally (fault) bounded by various basement-cored uplifts.
 |
West View of Western Bighorn Basin at Cody and Rattlesnake-Cedar Mountains
Verdant Cody sprawls in the foreground, abreast of the Shoshone River and Highway 14-16-20 that pass through town. The NW-SE ridge is formed by Rattlesnake Mountain and the north and Cedar on the south. The two summits form a massive anticline along the western boundary of Bighorn Basin. In the distance rises snow-capped summits of the Absaroka Volcanic Range with Yellowstone (unseen) lying beyond. Go there: 44°31′24″N 109°3′26″W
|
THE INTERMONTANE BIGHORN BASIN
The basin's alluvial sediments, as much as 25,000 feet in depth, are derived from the basement arches or uplifts that border the basin. Snow-capped, rugged summits and above 13,000 feet, the Pryor and Bighorn Mountain Ranges lie on the north and east, and the Bridger, Owl, Absaroka and Beartooth Mountains are on the south and west. Bighorn Basin drains to the north by tributaries of north-flowing Bighorn River such as the Shoshone that enters the basin between Rattlesnake and Cedar Mountains to the west of Cody.
Wyoming's basin and uplifts are not to be confused with the extensional landscape of the Basin and Range Province in Nevada and Mexico, which is associated with the Farallon's demise beneath North America. Basins such as the Bighorn are products of Laramide flat-slab subduction and thick-skinned deformation during Cretaceous-Paleogene compression. They possess an Archean basement in Wyoming, which in the basins occurs at depth, being down-warped, and in the ranges more surficially, being up-warped.
 |
Bighorn Basin
Modified NASA Satellite View |
STINKING WATER RIVER
Around 1807, so the story goes, Lewis and Clark expedition veteran and later solitary explorer and mountain man John Colter visited the as yet unnamed Cody region and crossed Stinking Water river at the location of a major 'Indian trail' ford. The river was named as such for good reason by the regional Crow Native Americans, who call themselves the Absaroka tribe. It's thought that the Crow, Shoshone, Bannocks and Nez Percé journeyed east over the Absaroka and Beartooth mountains to hunt the plains buffalo and test heralded medicinal values of "stinking waters” baths around Cody.
 |
Cody from the Southeast
Rattlesnake and Cedar Mountains form an imposing ridge across the skyline to the west of Cody (pop. ~500). Geologically enigmatic and highly debated Heart Mountain lies to its north (far right) and the gap to the south (far left). From USGS and Fisher, 1906. |
Understandably, the town's residents renamed the river around the turn of the century for obvious reasons, since things were starting to happen in Cody Country. By now, it had a new reservoir and dam built for purposes of agriculture and had a spur from Montana extended by the Chicago, Burlington and Quincy Railroad.
With federal money pouring in, the town was beginning to prosper and grow, and Buffalo Bill seized the opportunity to take advantage of it. He built the Irma Hotel in 1902. The "Grand Old Lady of Cody", named after his youngest daughter, still stands on the town's main street. Everyone from Colonel Cody, whose personality and fame kept the place packed, to princes and potentates, Indian chiefs and cowboys “came as they were” to the Irma.
 |
Northwest Wyoming in 1897 Rand McNally Map
The town of Cody, the Rattlesnake-Cedar Mountain complex, the roadcut with the Great Unconformity and the region of the future Buffalo Bill Reservoir at the confluence of North and South Forks lie within the circle. Also note Stinking Water River (labelled as such) drained the western Bighorn Basin. Modified from image at mapgeeks.org
|
COLTER'S HELL
Colter described the area west and southwest of Cody as having a "boiling tar spring" (submerged beneath Buffalo Bill Reservoir), fumaroles (vents of steam and sulfur), numerous sink holes and now-parched travertine cones that he reported to be geysered. The odoriferous river undoubtedly reminded him of nearby Yellowstone, which he had visited, and was later applied to the PArk's geyser area. It begs the question whether their hydrothermal geologies are related.
 |
DeMaris Vents of Colter's Hell
Along the Shoshone in Cody West of town and delineated by Wyoming Historical Marker, the vents are on the banks of the Shoshone on the north side of 14-16-20. The red strata of the Triassic Chugwater Formation conforms to the dip of the submerged eastern limb of Rattlesnake Mountain anticline. White travertine deposits lie along the banks and eroded ledges above the river. Tilted oil-producing, Permian Phosphoria Formation strata are visible in the banks. Sinkholes and travertine cones dot the landscape. Go there: 44°30'42.48"N, 109° 6'58.30"W
|
The region's hellaceous name was likely penned by local mountain men and fur trappers. Yet surprisingly, geothermal activity at Colter's Hell is unrelated to Yellowstone. Snowmelt and rainfall hydro-charge Absaroka aquifers to the east of Yellowstone. Flowing down gradient through faults and fractures in the crust, the waters reach sufficient depth, possibly near shallow magma bodies, to heat in excess of 43°C (110°F).
Rising along faults largely on the east flank of mostly Cedar Mountain, hydrothermal water discharges from vents and hot springs in the oil-rich, Permian Phosphoria Formation. Along the Shoshone River west of Cody, active and paleo-hot springs deposit travertine (precipitated calcium carbonate derived from underlying Mississippian Madison or possibly Ordovician Bighorn limestones) and sulfur (also associated with host limestone strata). The process is reminiscent of nearby Mammoth Hot Springs in Yellowstone near the Montana line.
 |
Hydrothermal Vents and Mineral Deposits along the Shoshone River
For four days in March, 2020 near Cody, vents spewed noxious sulfur gas with redeposition of travertine and sulfur along its banks that formed between the Gypsum Spring and Sundance Formations. Geologically famous Heart Mountain lies on the horizon. Photograph of Dewey Vanderfoff from BillingsGazette.com. Go there: 44°31'3.18"N, 109° 5'49.15"W
|
RATTLESNAKE AND CEDAR MOUNTAINS
Back on the highway, we're heading to the gap between Rattlesnake and Cedar. We're on the subtly upturned, synclinal shoulder of downwarped Bighorn Basin that succumbed under Laramide compression. A number of river terraces form benches around us that overlie down-dipping strata on Rattlesnake's hindlimb with an angular unconformity.
To the north, remnants of Cretaceous strata from the Western Interior Seaway form a series of erosion-resistant, sandstone hogback ridges that poke through eroded, softer shales and a blanket of Quaternary colluvium shed from the ridge to the west and alluvium from the meandering Shoshone.
 |
Northwest View of Rattlesnake Anticline
Some 25 km-long and NW-striking, Rattlesnake and Cedar Mountains form an imposing ridge between Cody on the east and the Absaroka Range and Buffalo Bill Reservoir immediately on the west. Shoshone Canyon and River occupies the gap between the two edifices. Note the location of the Great Unconformity, the Paleozoic strata that blankets the anticline and Cretaceous strata on the lowest limbs. |
Hematite-red patches of Chugwater sandstones capped by whitish gypsum off to the north were delivered during the Triassic by the regressing coastal sea and fluvial and lacustrine (lake) environments. It's the same formation we spotted in town in the bend in the Shoshone only tilted. As we head further upshoulder, we're travelling back in time through deposits of the eastern epeiric sea that will transition to marine deposition that came from the west.
LARAMIDE ANTICLINES
Imposing Rattlesnake-Cedar Mountain ridge is a landform called an anticline - an upward fold in the strata that slopes bi-laterally downward from the crest. Rattlesnake anticline and the Bighorn synclinal basin formed near the end of the Cretaceous when the landscape succumbed to Laramide compression and wrinkled like a rug squeezed together from the sides.
As a result, Paleozoic and Cretaceous strata that unconformably rests on an Archean basement bowed up, as the reverse occurred in the down-bowing basin that gradually filled with sediment. It left the Great Unconformity deeply buried in the basin but more shallow (and accessible to viewing) within the anticline. In time, uplift of the anticline unroofed the Mesozoic strata and left patchy remnants and erosion-resistant hogbacks on the lowest limbs.
 |
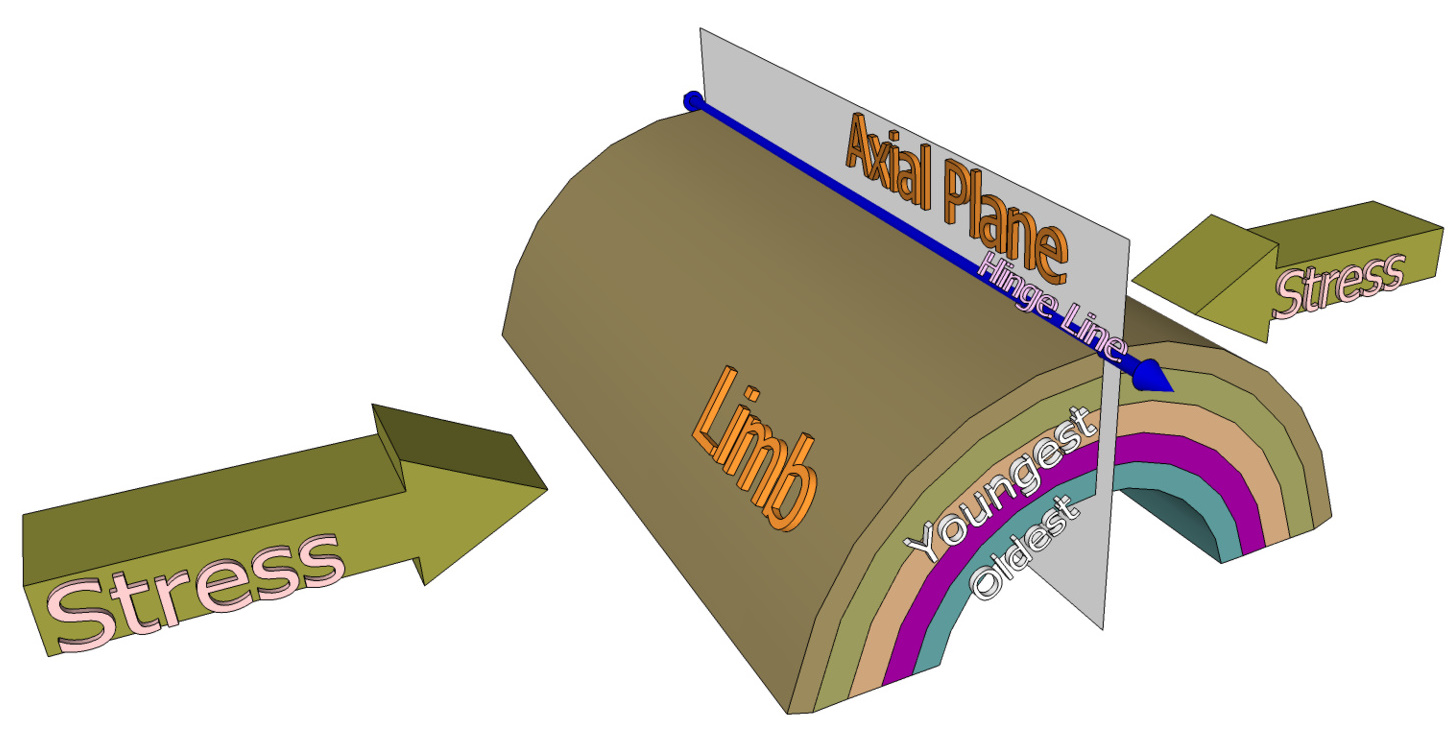
Diagram of an Idealized Anticline
The reverse of a syncline, oldest beds are at the uplifted center and younger are toward the exterior of the convex upfolded arch of strata. An axial plane transects the crest where curvature is greatest at the hinge zone, although, depending on compressive forces, the anticline may be asymmetrical. With relief, youngest strata erosively unroofs from the crest and flanks called limbs. Ridge-shaped anticlines typically develop above thrust faults during crustal deformation. Overturned anticlines are asymmetric with a limb that has been tilted beyond perpendicular and beds in the limb that have flipped over. The ends may be single or double-plunging. From Wikiwand.com |
PALEOZOIC STRATA ON DISPLAY
From a distance in the walls of the gap, we can see once-horizontal, now gently tilted Paleozoic strata from the Cambrian through Permian. And, as we begin to climb the asymmetric anticline's eastern hindlimb, we encounter the Phosphoria Formation, so-named for its marine-derived phosphates. Each formation tells a geologic story of the involving western continent and conditions as it was flooded by rising and falling seas from the west.
For example, the Phosphoria's limestones, shales and sandstones formed in the Permian when epicontinental seas (on shelves and shallow interiors versus deep marine basins) regressed from the sub-ranges of the long-gone Ancestral Rocky Mountains. Trapped hydrocarbons in the stratum have become an important source of oil and gas in Bighorn Basin, while phosphorus for fertilizer became concentrated from evaporating sea water.
 |
Heading into the Gap from Cody on Highway 14-16-20
Initially deposited horizontally (according to one of Steno's laws of stratigraphy), down-folded Paleozoic strata of the backlimb (plunging flank of the anticline) tilts to the east within the notch. Cody's Yellowstone Regional Airport is built on graded Quaternary alluvium derived from the upfolded Laramide antiform and others in the region.
|
Further west, we encounter progressively older, anticline-tilted marine strata of Pennsylvanian Tensleep dolomites, Mississippi-Pennsylvanian Amsden shales and sandstones and Madison limestones (Redwall equivalent in Grand Canyon). A time of uplift and erosion, no rocks of Silurian age are known in Bighorn Basin or Wyoming. Further back in time, transgressive deposition resumed with Devonian dolomites of the Three Forks and Jefferson Formation and Ordovician Bighorn Dolomite.
A TRANSGRESSIVE SEQUENCE
Entering the gap, we've crossed over the Shoshone River and arrived within the Cambrian core of Rattlesnake anticline. We're on Laurentia's continental shelf that stretched the length of western North America's coastline from north to south. It's a classic, time-progressive, quiet-water, marginal marine succession that's equivalent to the Grand Canyon's Tonto Group.
The sequence consists of lowermost deep-water sands, intermediary muds and uppermost, shallow-water carbonates deposited landward as Cambrian sea level gradually rose. Preserved and lithified, they form the sandstones, shales and limestones that lie above the Great Unconformity both in the Grand Canyon and here in the gap.
 |
Cross-Section of an Idealized Marine Trangressive Sequence onto a Continental Shelf
Modified from marlimiller.com
|
THE CAMBRIAN SEQUENCE
The thickness of the transgressive succession at any given point is dependent both on one's east-west position, since marine waters flooded from the west, and due to topography of the underlying Precambrian basement below the Great Unconformity.
From top to bottom, the Tonto-like sequence consists of Late Cambrian offshore glauconitic (greenish sandy, micaceous) limestones and shales of the Gallatin Formation that overly the Middle to Late Cambrian Gros Ventre Formation with the Middle Cambrian Flathead Sandstone that forms the base of the tripartite succession.
 |
North View of the Hindlimb of Rattlesnake Anticline within the Gap
The gently sloping, east hindlimb of Rattlesnake anticline is clearly visible. Higher up on the flanks of Rattlesnake Mountain are middle and late Paleozoic marine strata. At the bottom of the Paleozoic succession and tilted in concert with the hlindlimb, the Great Uncomfornity is exposed in the roadcut between thickly-bedded strata of Flathead Sandstone and above Northwest Wyoming's Archean basement.
|
GEO-ACCOLADES OF THE FLATHEAD
Up to 300 feet thick, its a reddish-brown to pale-orange and yellowish-gray, coarse to fine, cross- and parallel-bedded arkosic sandstone (mica, quartz and feldspar-rich). It's the oldest and basalmost sedimentary strata in Wyoming and the first-to-form Paleozoic strata and Sauk sequence of western North America. It's also the equivalent of the Tapeats Formation in the Grand Canyon and unconformably resides over the region's Archean igneous and metamorphic rocks with the temporal gap of the Great Unconformity interposed.
Crystalline highs were gradually flooded islands, awash in the Cambrian sea. Early in its formation, ephemeral and ever-changing, braided streams and fluvial systems (below) left an indelible, lens-shaped mark as the sea increasingly flooded the landscape.
 |
The Slabby, Ledgey Flathead Sandstone
A planar-laminated lens-shaped stream channel appears in cross-section within the exposure lying below a cross-bedded, rapid facies change and is more sub-angular conglomeritic and feldspathic than later in the formation. Being poorly-sorted, an occasional Archean gneissic fragment is incorporated within the sandstone matrix. The vertical cuts were created during construction of the roadcut and highway.
|
THE SAUK SEQUENCE OF THE WYOMING SHORELINE
The early fluvial systems (photo above) that delivered minerals seaward from topographic highs were remixed and redeposited by the rising sea. Age and paleocurrent indicators assign provenance of the Flathead's sands from Wyoming's Archean Beartooth-Bighorn Magmatic basement to the north (dotted blue arrow), while Tapeats minerals are from southerly Proterozoic Yavapai-Mazatzal granites (solid arrow).
In both cases, the sources are west of the Transcontinental Arch that stretches from the southwestern U.S. to south-central Ontario. Long gone, the prominent uplift was an extension of the Archean-age Canadian Shield of Laurentia's lower Paleozoic craton. The topographic barrier influenced early sediment transport patterns in the mid-continent of Laurentia from the Late Proterozoic and early Paleozoic by preventing southern and eastern Grenville minerals from reaching Wyoming and northern Arizona.
 |
Paleographic Middle Cambrian Reconstruction of Wyoming and Environs
Around 520 Ma, the Sauk seas transgressed over western Laurentia from west to east and deposited the Flathead Sandstone, known as the Tapeats in northern Arizona. Brown shaded rocks are Archean basement. Blue arrows reflect potential zirconic mineral sources via south-flowing streams from the Beartooth-Bighorn Magamtic Zone or north-flowing streams from the Transcontinental Arch. From Malone, 2017.
|
PALEOENVIRONMENT OF THE FLATHEAD
Its unassuming sands contain evidence of abundant benthic (bottom-dwelling versus pelagic or open water) marine assemblages preserved as worm trails and trackways. The lifeforms are thought to have been lobopodians, ancestors of modern onychophorons. The soft-bodied worm burrowed vertically into the substrate at a time in evolution when rapid development of seafloor biota and diverse body plans were beginning to occur.
In the earliest Cambrian, seafloor traces and tracks were unbranched, small, simple and oriented horizontally close to the sediment-water interface. But by the Middle Cambrian, deeply vertical-burrowing lifeforms were bioturbating (disrupt) sediments below the interface.
It was a monumental event that changed the world forever - when latest Proterozoic Ediacaran fauna (largely sessile, soft-bodied, tubular and frond-like organisms) explosively transitioned to Burgess biota that underwent a major taxonomic biodiversification and adaptive radiation.
 |
Heavily Bioturbated Tapeats at Plateau Point in Grand Canyon
The Tapeats and Flathead Sandstones were likely hosts to the same biota as the famous Burgess Shale in western Canada. Typically, biogenic facies include skolithos (simple, vertically dwelling tube) and cruziana (feeding locomotion trace), which as |
THE PRECAMBRIAN-CAMBRIAN BOUNDARY
The deep burrowing that changed the seafloor marked an important time in Earth history, above which modern ecological systems began to evolve. Soon after, the Middle Cambrian rock record became crowded with the fossils of animals that possessed scales, shells and platy armor. The world's first predators had arrived!
Exposed horizons of Flathead Sandstone reflect this change, albeit subtly, and are part of the mystery and awe that is associated with the Great Unconformity. What global geological events are thought to have led to the formation of the temporal gap? See my post Part here for some of the answers.
 |
Changes in Seafloor Biota across the Precambrian-Cambrian Boundary of the Great Unconformity
Modified from The Rise of Animals by Mikhail A. Fedonkin et al, 2007.
|
THE GREAT UNCONFORMITY AT SHOSHONE CANYON
Our westward travel into the gap has finally brought us to the Great Unconformity at the core of Rattlesnake anticline. The temporal gap formed in the Middle Cambrian with the Flathead's deposition over the crystalline foundation of the Beartooth-Bighorn Magmatic Zone.
Tilt of the strata reflects the gentle dip of the Rattlesnake anticline's eastern hindlimb. A dramatically exposed, steep forelimb lies past the third tunnel through the anticline The temporally 'greater' Great Unconformity exceeds the duration of the contact in Grand Canyon by over a billion years, again owing to the extreme age of the Northwest Wyoming Province.
 |
Julia daintily perched on the Great Unconformity
Higher in the four-member, beach to offshore Flathead sequence bedding appears thicker and increasingly interbedded and cross-bedded perhaps as longitudinal bars migrated their locale.
|
SHOSHONE CANYON
A visit to the Great Unconformity wouldn't be complete without peering into Shoshone Canyon and River and travelling through the west-east gap and tunnels to Buffalo Bill Reservoir and Dam to the west. The ~5 km-long abyss occupies the gap between Rattlesnake Mountain on the north from smaller Cedar Mountain (formerly called Spirit Mountain) on the south.
A vertiginous glimpse of Shoshone Canyon and River can be had at the end of a south to the south before the first tunnel. It provides a wonderful view of the Archean concoction of rocks that lie immediately below the Great Unconformity - Beartooth-Bighorn's igneous and metamorphosed plutonic rocks (gneiss, schists and amphibolites) and injected diabase (intrusive basalt) dikes.
 |
The Archean-age Crystalline Walls of Shoshone Canyon and Greenish Shoshone River
It's appearance below the Great Unconformity has a familiarity that can be globally observed. |
CODY ROAD - OLD AND NEW
At the time Cody was founded, the only practical access to Yellowstone was from the north via Montana by pack train. An alternative but arduous route was around Cedar Mountain and followed the South to the North Fork of the Shoshone to Yellowstone on rugged trails and a primitive wagon-track. Once federally funded, Shoshone Dam and its treacherous and oft-flooded service road along the river changed that.
Beginning about 1906, the Cody Road began to evolve into today's Buffalo Bill Scenic Highway (aka "Yellowstone Road"). "The most scenic fifty miles in America" according to President Theodore Roosevelt reduced travel between Cody and the park from days to 45 minutes, which became suitable for motor cars by 1916. On a grandeur scale, it's a section of U.S. Highway 14-16-20 that runs across Wyoming.
 |
Shoshone Canyon Facing West and the 'old' Cody Road
With permission of Mark Fisher from GeoWyo.com (here).
|
In anticipation of a heavily traveled road, Buffalo Bill built the Pashaka Tepee hotel near the entrance to Yellowstone and other lodges followed suit. In addition, there are a number of interesting geologic sites to visit along the way such as the Chinese Wall dike that emanates from Sunrise Volcano and the Holy City of volcaniclastic hoodoos.
 |
Buffalo Bill on the Porch of the Pashaka Tepee c.1905
From CenteroftheWest.com |
SHOSHONE RIVER
Farallon compressional tectonics conferred the landscape of Northwest Wyoming with a domal shape. During the Miocene, Basin and Range extension caused regional uplift that rejuvenated rivers such as the Shoshone, triggering erosional activities. And, being east of the hydrologic Continental Divide that crosses north to south through Yellowstone, rivers such as the Shoshone that drain the uplifts converged and flowed radially from the region.
As a result, like other rivers and streams in Bighorn Basin, the Shoshone travels WNW some 100 km across the relatively undeformed, downwarped depression, through and around rolling plains, flat mesas, badlands, benches, foothills and mountain ranges. Eventually, it merged with progressively larger rivers Bighorn, Yellowstone, Missouri and finally the Mississippi.
 |
Major Tributaries and Rivers that Drain Bighorn Basin
The Great Unconformity at Rattlesnake anticline is located at the red arrow just west of Cody. Notice the North and South Forks that headwater in the Absaroka Range and converge at Buffalo Bill Reservoir. |
HEADWATERS OF THE SHOSHONE
The 100 mile-long river originates within the rugged Pleistocene-glaciated, fluvially-reworked Absaroka Mountain Range. It's a 9,000 sq mi sub-range of the Middle Rockies that forms Yellowstone's eastern boundary and skyline. Heading due east, Shoshone's North and South Forks converge on the Buffalo Bill Reservoir that is impounded by Buffalo Bill Dam some six miles west of Cody. The road we're on follows the North Fork to Yellowstone's East Gate.
The waters are released from the dam into the Shoshone River that carved narrow Shoshone Canyon between Rattlesnake and Cedar Mountains and, over time, the gap between the two mountains. More on that later. This is the region of the 3,000 acre Buffalo Bill State Park named after Colonel William F. Cody, who owned land in these parts. We'll get to him later too.
 |
West Aerial View of "Cody Country"
With Yellowstone on the horizon, out of view beyond the Absaroka Range, the North and South Forks converge at Buffalo Bill Reservoir. Shoshone Canyon extends between Rattlesnake and Cedar Mountains and begins a serpentine course through Cody and across Bighorn Basin. The approximate location of the exposure of the Great Unconformity is delineated. |
A FEW WORDS ABOUT THE ABSAROKAS
It consists of 13 stratovolcanic centers composed of interlayered basalt (and some andesitic) lavas, and volcaniclastic rocks with lahar (volcanic mudflows) and tuffs (compacted and lithified ash-flows) that emplaced in a relatively short-lived but intense event in the Middle Eocene (~53-49 Ma). The volcanic province likely unconformably overlies folded and faulted Paleozoic, Mesozoic and lower Tertiary sedimentary rocks that, as mentioned, partially blanket Rattlesnake and Cedar Mountains and Bighorn Basin, but the difference ends there.
 |
South View of the Absaroka Range from the Beartooth Range in South-Central Montana
From Beartooth Pass at 10,947 feet, the Absaroka range has been extensively eroded with wind, water and ice since their volcanological inception some 50 million years ago. We're standing in the Beartooth Range across the Wyoming state line in Montana with a similar compositional basement of Archean age but uplifted during teh Laramide orogeny. The Absarokas form the boundary at Yellowstone's Eastern Entrance. Go there: 44°58.1′N, 109°27.9′W |
GEOGRAPHICALLY RELATED BUT GENETICALLY DISTINCT
Rattlesnake and Cedar Mountains form the eastern boundary of the Absaroka Range and are often geographically included with it, however, they differ in rock composition, structure, stratigraphy and provenance. Explanations for emplacement of the Absaroka Volcanic Province regard the southwestern edge of the demised Kula plate that subducted beneath the Pacific Northwest directly before arrival of the Farallon plate.
Two major Absaroka genetic theories include extension and accompanying upwelling asthneospheric (upper crust) magmatism that was produced at the Farallon-Kula spreading-ridge. Alternately, Kula plate roll-back that opened a window (or two) allowed ascending magma to initiate a shallow crustal melt in the 'non-arc' inter-plate interlude before being shut-off by incoming Farallon arc-related volcanics.
In contrast, genetically distinct Rattlesnake and Cedar Mountains succumbed to low-angle Farallon compression in the Late Cretaceous to early Eocene by folding and unroofing.
 |
Holy City along the North Fork
A massive volcaniclastic apron from the Absaroka's Sunlight Volcano left a chaotic, radiating array of layered mudflows, breccia lahars, lavaflows, dikes and tuffs. Visible from the interstate, volcaniclastic sedimentary rocks of the Holy City weathered into fanciful hoodoos. Remnants of Eocene-age petrified wood, preserved by rapid mudflow burial and silicification, can be found in gravel bars along the Shoshone's North Fork. |
Incidentally, similar volcanic belts and fields to the Absarokas are in eastern Washington, British Columbia, Idaho, Wyoming (Devil's Tower) and the Black Hills of South Dakota that emplaced from SW to NE in concert with Kula subduction. In addition, early geologists associated the Eocene Absaroka volcanics with Yellowstone caldera - the largest volcanic system on the North American continent - but it formed more recently in the Pliocene and Quaternary with a hotspot mantle plume genesis that is controversial and actively debated.
A BASEMENT-CORED LANDFORM
Rattlesnake and Cedar Mountains form the Rattlesnake Mountain anticline along with Horse Center anticline, a smaller, eroded extension to the south. Following the period of thin-skinned Sevier tectonics, Laramide compression not only uplifted the ranges that surround downdropped Bighorn Basin but, reactivating inherited (pre-existing) basement faults, folded the basin's Phanerozoic strata into a series of fault-related anticlines around its margins.
Wrinkling like a rug compressed together from the sides, it formed an elongated, A-shaped arch of Archean basement crust (Ag) blanketed with folded, once-horizontal sedimentary strata. In the case of Rattlesnake, compression from the west and antithetic (opposite-dipping and directed backward) backthrusts on the east that occurred as Bighorn Basin propagated to the east up-folded the anticline.
 |
North View of "Cody Country"
Defining and deciphering the tectonic events that contributed to basement fractures is challenging, since they may be related to the reactivation of inherited basement faults during Laramide compression and even subsequent Basin and Range extension that followed.
|
ANATOMY OF RATTLESNAKE ANTICLINE
It's a classic landform formed from Laramide compression. The west-facing flank or forelimb rises steeply to the crest or hinge-zone and descends gently on the east-facing a hindlimb. The anticline is completely draped with a massive fold of layered Paleozoic strata from which Mesozoic strata assumedly has unroofed.
As a result, progressively older Paleozoic strata is encountered with depth until Archean crystalline rock is reached at the core of the anticline (the reverse of synclines). As a result, facilitated by carving of Shoshone Canyon between Rattlesnake and Cedar Mountains, the Great Unconformity in the roadcut is exposed on the south face of Rattlesnake Mountain where the descending hindlimb is nearly horizontal on the eastern edge of Bighorn Basin.
 |
Idealized Anticline Diagram
From Speleothem of Wikimedia Commons |
THE BIG PICTURE
Marine sequences formed on the western passive margin of Laurentia as Pangaea began to assemble in the Paleozoic. At that time, the region of Bighorn Basin was part of the stable continental shelf and intermittently submerged. In the Pennsylvanian-Permian, it was drowned and uplifted as the Ancestral Rocky Mountain Range rose and was beveled by erosion.
During the Cretaceous, the part of Wyoming that is now the Bighorn Basin was near the western edge of the Rocky Mountain foreland basin, an elongate north-south structural depression that developed east of the tectonically active Western Cordilleran highlands. Throughout much of its history, the subsiding basin (a Farallon compressional, downwarped landform) was flooded by rising eustatic (global) seas that formed the broad Western Interior Seaway from the Arctic Ocean to the Gulf of Mexico, at its greatest extent.
The epeiric sea periodically delivered a complex pattern of intertonguing muds and sands to Northwest Wyoming but withdrew with the first impulses of the Laramide orogenic phase. Mesozoic rocks eroded off the anticline but appear on the shoulder of Bighorn Basin on RMA's lowest flanks (JTu and Ku), where not blanketed by Quaternary alluvium.
 |
Generalized Bedrock Geologic Map of "Cody Country"
The Eocene-age volcanic Absaroka Range dominants the landscape to the west of the Laramide compression-formed Rattlesnake Mountain anticline.
Modified from Wyoming State Geological Survey
|
ANATOMY OF AN ANTICLINE
Basement-cored, Laramide compression at Rattlesnake Mountain anticline (map below) produced a horizontally-stacked, elongated arch of Paleozoic sedimentary rocks with an underlying foundation of Archean rock. The anticline's forelimb steeply upfolds to the crest, which is assymetrically positioned to the west, drapes across the hingeline (inflection point) and gently downfolds on the backlimb.
As a result, progressively older strata is encountered within the anticline's core, which is the inverse of a syncline. On our drive from east to west between Rattlesnake and Cedar we witnessed the anticline's stratal composition. and structure. Furthermore, around Bighorn Basin, compressional and antithetic (opposite-directed) back-thrusts deformed originally flat-lying Paleozoic and unroofed Mesozoic sedimentary successions into a series of synclines and anticlines.
 |
Simplified Geologic Map of Rattlesnake Mountain Anticline
With a steep fore limb and gently sloping hindlimb, Cretaceous strata has been unroofed across the folded Paleozoic sequences. Shoshone Canyon has excavated to the Archean basement as it slices through the anticline that's oriented NW-SE on the landscape. A cross-section of Laramide landform is represented below along an A-A' transect. Modified from Beaudoin, 2012.
|
BUFFALO BILL RESERVOIR
By now, we've traveled beneath Rattlesnake Mountain some six miles west of Cody and emerged from the third tunnel, the longest in the state. West of the gap on the north side of the reservoir, the landscape dramatically opens up into an expanse occupied by the 8,315 acre Buffalo Bill Reservoir. It's heart-shaped by convergence of the North and South Forks on the west with Sheep Mountain in between, a geological enigma in its own right.
 |
Confluence of North and South Forks in the Region of the Future Reservoir around 1906
From WyomingTrailsandTails.com |
The reservoir lies within Buffalo Bill State Park established in 1957, was named for William Frederick Cody, best known as "Buffalo Bill", who owned land in the region in the 1870s. In addition to founding the eponymous town in 1896, he envisioned and was influential in bringing irrigation and agricultural development to Cody and some 107,000 acres of the northwestern Bighorn Basin's semi-arid pasture and farmland.
 |
Southwest view of Buffalo Bill Reservoir and Dam, Shoshone Canyon and Gap
The U.S. Reclamation Service that conducted the irrigation project required sacrificing settled ranch land along the North and South Forks and flooding of the small community of Marquette. The government bought the properties for roughly $400,000 in 1905. |
"BUFFALO BILL"
I would be remiss without further mention of Colonel (an appointed political title) Cody (1846-1917 1917). Born in Iowa, he was a flamboyant, true-to-life living legend, showman and entrepreneur of the Old West. His colorful and varied life included jobs as a ranch hand, wrangler, wagon train driver, gold prospector, fur trapper, Pony Express rider, Civil War veteran, civilian scout and Medal of Honor recipient buffalo hunter.
As if not enough, he's best known for his cowboy, circus-like, open-air, travelling Wild West Show that entertained and delighted common folk audiences, celebrities, dignitaries and royalty in the U.S., Great Britain and continental Europe for over 10 years beginning in 1883. It celebrated America's uniqueness and offered a romanticized version of the Old West.
 |
Distinguished Portrait of Buffalo Bill from 1911
From the U.S. Library of Congress in the Public Domain |
The action packed show featured rifle sharpshooter Annie Oakley and an array of western performers doing rope tricks, chasing live buffaloes and galloping about while enacting mail delivery of the Pony Express, the Battle of the Bighorn of Custer's Last Stand and action packed stagecoach holdups with rifles and pistols blazing. Its Congress of Rough Riders of the World included cavalries, Native Americans and horsemen in full regalia from the West, South America, Turkey, Arabia and Mongolia. It must have been a sight to see, hear and smell!
A must-see while in Cody is the world-class Buffalo Bill Center of the West. It's five award-winning, expansive museums under one roof include - the Plains Indians, Whitney Western Art Museum, Draper Natural History Museum, Cody Firearms Museum and of course the Buffalo Bill Museum. There are countless interactive exhibits and live animal exhibitions.
 |
| Poster of Buffalo's Bills Wild West Shows from 1908 |
BUFFALO BILL DAM
Buffalo Bill joined John Wesley Powell, famous explorer of the American West and Grand Canyon, and others in the late 1800s to implore Congress to create an energy agency dedicated to developing water systems out West. As a result, the dam was built between 1905 and 1910 by the Bureau of Reclamation below the convergence of the North and South Forks of the Shoshone across the narrowest part of Shoshone Canyon on the west side of the gap.
Originally called Shoshone Dam, it was renamed for the region's namesake in 1946. At 325 feet high, it was a landmark engineering achievement with accolades that included highest concrete dam in the world, listing on the National Register of Historic Places, declaration as a National Civil Engineering Landmark and prototype for the Hoover Dam completed in 1936. Read about it
here.
 |
Julia atop Buffalo Bill Dam
|
Facing west from the dam , the greenish reservoir is embraced by forelimbs of Rattlesnake (right) and Cedar Mountains (left) comprised of west-dipping limestone strata. In the distance, between converging North and South Forks lies prominent Sheep Mountain, and on the horizon are summits of the Absaroka Range with Yellowstone beyond view.
Sheep's history mimics that of long-enigmatic Heart Mountain just north of Cody. In the Middle Eocene, fed by gravity and possibly thermally-triggered by nearby Absaroka volcanic activity, Heart Mountain formed in the Eocene, when a massive block of Ordovician Bighorn Dolomite - the size of Rhode Island - catastrophically slid some 50 km along a low-angle detachment fault and came to lie allochthonously over Eocene Absaroka volcanics. It's the largest known terrestrial landslide that remains the subject of geological debate.
 |
West View of Buffalo Bill Reservoir from atop Buffalo Bill Dam
|
Buffalo Bill Dam is classified as a constant radius, concrete (versus masonry used at the time) arch-gravity dam that curves upstream to apply water pressure to the narrow canyon walls. Constructed 68 feet below the bed of the river, the dam was raised another 25 feet in 1993 to increase the reservoir's capacity to its present 646,563 acre-feet of water. Two long outlet tunnels carry water to cropland of western Bighorn Basin. Beginning in 1920, an interconnecting system of hydroelectric power plants was developed.
 |
Dizzying View into the Narrowest Aspect of Shoshone Canyon
The dam's spillway consists of a rock weir, an intake channel and a tunnel. It allows the controlled release of water into the Shoshone River. An original powerplant , addition 1992 |
THE ARCHEAN CRATON OF NORTH AMERICA
Facing east from the dam, the geology of the Archean-age granite and granitic gneiss of Shoshone Canyon is on glorious display. High above the chasm, overlying and tilted Paleozoic strata of Rattlesnake Mountain are visible on the anticline's descending hindlimb.
Evidence of reverse, hanging-wall backthrusting, previously mentioned, is exposed in the canyon walls between steeply dipping basement rocks and Cambrian to Ordovician rocks of the anticlnal forelimb, having been Laramide deformed coevally.
 |
East View of Shoshone Canyon from atop Buffalo Bill Dam
The canyon service road (dotted arrow) was built during construction of the dam. A small section of Highway 14/16/20 can be seen emerging from the second of three tunnels (solid) cut into the south face of Rattlesnake Mountain. Due to upward inclination of the anticlinal hindlimb, the Great Unconformity lies above the tunnel. Higher in the gap, middle to late Paleozoic sedimentary strata likewise dip to the east.
|
FORELIMB OF RATTLESNAKE ANTICLINE
From the parking area, the view to the north reveals the stratigraphy that constitutes the steeply descending limb of the anticline. Thin and thickly bedded, tilted Paleozoic marine sandstones, shales and limestones delivered formed in marine waters of the proto-Pacific from mid-Cambrian to Pennsylvanian time, can be seen rising toward the hingeline.
It has been unroofed of late Paleozoic marine strata and likely from the Mesozoic as well, the latter delivered from the long-gone Western Interior Seaway on the east. Unseen, the Great Unconformity is buried below the Middle Cambrian Flathead Sandstone at the parking area. The geologic feature formed only after an incredibly long hiatus of erosion, uplift and denudation of the Archean basement before being blanketed by the deposition of Cambrian-age strata.
 |
Overturned Forelimb of the Western Rattlesnake Anticline
|
LARAMIDE BASEMENT-CORED FOLDING, THRUST BELTS AND REVERSE FAULTS
The contractional event that resulted in the formation of Rattlesnake anticline involved Archean basement structures and some 7 km of overlying strata from the Paleozoic and Mesozoic. Steeply-overturned strata confirm our position west of the crest of the anticline (parking area at arrow below).
We are within the region of an overturned forelimb. It's a decoupled basement wedge that formed by multiple superficial and deep, reverse fault splays that are branching faults near the termination of the major fault. As a result, displacement has been spread over a large area over the western flank of the anticline. Theories of the mechanics of fault propagation abound as do the faults themselves (See references).
 |
Fold Geometries and Stratal Relationship of Rattlesnake Anticline Facing North
Thrust faults of the anticline's western aspect were generated during the Laramide phase (orogeny) of Farallon compression in the Late Cretaceous that folded the landform. Note the location of the observer facing north in the parking area in the above photo. Paleozoic forelimb strata is overturned and near vertical, whereas it's gently downfolded on the hindlimb. Cambrian strata are concordant across Shoshone Canyon on Cedar Mountain. Not all formations are indicated. Photo modified from summitpost.org. |
WHAT CAME FIRST? THE RIVER OR THE RIDGE
We've witnessed the Shoshone River flowing west to east through Shoshone Canyon between Rattlesnake and Cedar Mountains (dotted white line). Why did the river 'choose' to maintain such an albeit direct but laborious course through erosion-resistant crystalline rock rather than detour to the south around Cedar Mountain (red dotted line) through erosion-susceptible sedimentary rock at a lower elevation?
It begs the question what formed first on the landscape - the river or the ridge? In other words, is the course of the river older than the uplift of the mountain through which it has carved a canyon? Can river sediments or those shed from Rattlesnake be age-dated and provide clues to the conundrum? It's a question of antecedence or superposition (aka superimposition).
ANTECEDENT AND SUPERIMPOSED RIVERS
Antecedence describes a process whereby a river that flows across a subdued landscape predates the formation of an uplifting mountain and maintains its course, cutting a canyon in spite of resistant lithology and structure.
In contrast, superposition defines a condition where the river downcuts through erodable sediment to a preexisting, buried topographic landform and maintains its course, carving a canyon as the landform is gradually exhumed.
 |
Fluvial Processes of Antecedence and Superposition
In both cases, the river flows from right to left across a subdued landscape and maintains it s course.
With permission, image from Carving Grand Canyon by Wayne Ranney, 2012. See references. |
TWO POSSIBLE SCENARIOS WITH IDENTICAL OUTCOMES
Emplacement of the Absaroka Volcanic Field in the Middle Eocene and formation and uplift of the Rattlesnake anticline in the Cretaceous-Eocene time interval overlap, although the anticline likely preceded it, in addition to renewed uplift in the mid-Neogene. If the anticline had been buried beneath sediment or even partially exposed, the Shoshone could have established a course over it and maintained its course during its exhumation. Shoshone-delivered Absaroka rocks in river terraces around Cody support such an antecedent scenario.
On the other hand, if the Shoshone maintained it course as it eroded down to a sediment-buried Rattlesnake-Cedar Mountain range that had previously formed, a likely possibility, it would have maintained its course carving Shoshone Canyon through the anticline. Studies in the greater West support this type of superposition scenario. What's your genetic hypothesis? I cast my vote for superposition.
I HAD A BLAST THANKS TO MY INTREPID TRAVEL COMPANIONS
Science teacher, field biology aficionado, great photographer and daughter Julia Share
 |
Buffalo Bill Dam Ball Plug
Constructed of wood and concrete, it was one of two used to halt the flow of water through two 42-inch diameter conduits within the base of the dam that facilitated repairs and maintenance of downstream equipment. I'll use flash fill next time! |
Renown geologist, author, guide and good friend Wayne Ranney (
here)
 |
Buffalo Bill Center of the West in Cody
Me, Buffalo Bill and Wayne
Go there: 44°31'28.71"N, 109° 4'26.00"W |
By the way, the Whitney Western Art Museum of Cody's Buffalo Bill Center, that displays original paintings, sculptures and prints of the American West from the early nineteenth century, has two oils on canvas by William Tylee Ranney (1813-1857). From Connecticut, he's a distant relative of Wayne's best known for his genre paintings of everyday life on the frontier.
 |
"Advice on the Prairie"
In this painting, Ranney portrayed a group of Western immigrants that included a family camped with their wagon for the evening. They're listening to tales from a scout of what they might encounter on their journey. Oil on canvas. |
REFERENCES AND RECOMMENDED READING
• Ancient Landscapes of the Colorado Plateau by Ron Blakey and Wayne Ranney, Grand Canyon Association, 2012.
• Carving Grand Canyon - Evidence, Theories and Mystery, Second Edition by Wayne Ranney, 2012.
• Detrital Zircon Geochronology and Provenance of the Middle Cambrian Flathead
Sandstone, Park County, Wyoming by David Malone et al, Mountain Geologist, April 2017.
• Fracture Analysis of Circum-Bighorn Basin Anticlines, Wyoming, Montana by Julian Stahl, Montana State University Thesis, November, 2015.
• Geologic Map of the Cody 1°x2° Quadrangle, Northwestern Wyoming by William J. Pierce, 1997.
• How Rivers Get Across Mountains: Transverse Drainages by Phillip H. Larson, Anals of the American Association of Geographers, August 2016.
• Impact of Fracture Stratigraphy on the Paleo-hydrogeology of the Madison Limestone in Two Basement-involved Folds in the Bighorn Basin (Wyoming, USA) by Mickael Barbier et al, Tectonophysics, July, 2012
• Late Cretaceous–Early Eocene Laramide Uplift, Exhumation and Basin Subsidence in Wyoming: Crustal responses to Flat Slab Subduction by Majie Fan et al, Tectonophysics, March, 2014.
• Solid Mineral Occurrence and Development Potential Report - Bighorn Basin Resource Management Plan Revision Project by Bureau of Land Management Cody Field Office, Wyoming, 2009.
• Structural and Microstructural Evolution of the Rattlesnake Mountain Anticline: New Insights into the Sevier and Laramide Orogenic Stress Build-up in the Bighorn Basin by Nicolas Beaudoin et al, Tectonophysics, November 2012.
• Techniques for Uunderstanding Fold-and-Thrust Belt Kinematics and Thermal Evolution by Nadine McQuarrie et al, Geological Society of America, Memoir 213, 2017.
•
The Great Unconformity - Geology of Wyoming by Ken Steele et al, Geowyo.com (
here). •
The Wyoming Province: A Distinctive Archean Craton in Laurentian North America by Paul A. Mueller et al, Canadian Journal of Earth Sciences, 2006.
• Thick-skinned Tectonics and Basement-involved Fold-thrust Belts: Insights from Selected Cenozoic Orogens by Nicolas Bellahsen et al, Cambridge University Press, January 2016.